. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
M. Hildebrand, 1962:
"Further studies on locomotion of the cheetah",
Journal of Mammalogy
42, 84-91 (1962).
- . . . 2003).3
-
L.L. Marker and A.J. Dickman, 2003:
"Morphology, physical condition and growth of the cheetah
(Acinonyx jubatus jubatus)",
Journal of Mammalogy84, 840-850 (2003).
- . . . 2003):4
-
R.McN. Alexander, 2003:
Principles of Animal Locomotion (Princeton University Press, 2003).
- . . . \/.5
-
This seems pretty quick, but within reason.
A Thomson's gazelle reaches 15 m/s starting from rest
in approximately 5 seconds (Elliott
et al., 1977);
but the gazelle is more of a middle-distance runner,
whereas the cheetah is Nature's ultimate "power sprinter".
Assuming that the acceleration of the body and the foot are equal
may be an oversimplification.
If we throw in a factor of two
to give the average acceleration
between 8.6 m/s2 and zero,
we get t1 = 3.4 s,
which is still a little quicker than the gazelle
but a lot more challenging for the cheetah.
- . . . \/.6
-
This value is consistent with Kh = 270 J calculated for
just the hindlimb of a cheetah by
M. Hildebrand and J.P. Hurley, 1985:
"Energy of the oscillating legs of fast-moving
cheetah, pronghorn, jackrabbit and elephant",
Journal of Morphology184 23-31 (1985).
- . . . fuel.7
-
There is an assumption in the wording of the question
that the same force F is applied throughout,
even after the cheetah has reached top speed Ut
and ceased accelerating.
On one hand, as any sprinter can attest,
running at top speed is not effortless,
even though acceleration is finished;
there is friction to overcome,
and merely moving one's legs back and forth that fast
takes an enormous amount of work!
On the other hand, this does not translate into
an externally applied net force F.
However, if we approximate the net rate of work
as being the same (namely the maximum the animal can manage)
throughout the run, and we know that it mostly goes into
acceleration at the beginning, we can get a fair estimate
of the net energy expended by pretending that
the original F is exerted through the entire distance
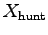 .
.
- . . . metabolism.8
-
T.M. Williams, G.P. Dobson, O. Mathieu-Costello, D. Morsbach,
M.B. Worley and J.A. Phillips, 1997:
"Skeletal muscle histology and biochemistry of an elite sprinter,
the African cheetah",
Journal of Comparative Physiology B167, 527-535 (1887).
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .